| Home | About | Helping Us | Contact | Mailing List |
| Features: |
|
Correa PN, Correa AN
Aurora Biophysics Research Institute
ABRI Monographs: Biophysics Research AS2-33
© Correa & Correa, 1979, 2006
All rights reserved.
Experimental proof is presented for the immediate physiological effect of human exposure to a "medical ORAC" on body temperature taken orally. The effect is objective, reproducible and weather dependent; it is always positive on anticyclonic days, plateaus at 15 to 30 min of exposure, is independent of time of day and tested subject. Time course studies indicate that the temperature elevation increases with time of exposure, up to 45 min, with significant elevations occurring as early as 5 min of exposure. The coefficient of temperature rise is steeper for start temperatures closer to the normal temperature floor (35.7-35.9°C) than to the normal temperature ceiling (37.1°C). At the 35.7°C starting level and at 15 min, the coefficient is 0.04°C/min, whereas starting 1.1°C higher, at 36.8°C and at 15 min, the coefficient is lower - at 0.02°C/min. A typical mean ΔT value for the observed induced functional hyperthermia at 15-30 min is 0.6°C, with typical errors (SEM) of 0.06 to 0.13°C, irrespective of time of day on anticyclonic days.
"Faraday cages" made of metal conductors were first invented by Michael Faraday to electrically shield an object of study: "Faraday built a cube twelve feet on an edge. Copper wire was passed over it in all directions until a mesh had been formed. Paper upon which tinfoil bands had been fastened was then used to cover the cube so that the whole was an excellent conductor. Faraday then entered the cube carrying instruments used for detecting electrostatic charge. When the cube was charged to such an extent that sparks flew off at the corners and the tinfoil strips stood straight out with brushes of electricity surrounding their ends, Faraday could not detect the slightest effect." [1]
Note that Faraday's original cage already contained a covering of 'organic' or dielectric material (in fact a semiconductor, since paper is a very good conductor for high-voltage currents), and was in essence a 1x or 1-ply ORAC enclosure. It was not, however, made of ferromagnetic material, nor a very good thermal and electric insulator.
Since that time, Faraday cages or rooms have become standards in the radio and recording industries - where it is important to shield an environment from electromagnetic signals and their noise contribution - as well as in electrophysiology and other areas of scientific research, where it matters to filter out electromagnetic noise from light fixtures, ordinary electric motors, and other electrical equipment .
In essence, this well-known property of metallic enclosures already indicates that a metal cage can block - and thus alter or transform - any incoming flux of charges, ambipolar radiation included. We have also shown that if the cage is made of ferromagnetic material, it can slow down or arrest the so-called "spontaneous leakage" of charge from electroscopes placed inside of it, irrespective of charge polarity [2-4].
In 1941, Wilhelm Reich created the orgone accumulator (ORAC) by wrapping a ferromagnetic Faraday cage with repeated alternating layers of insulator/dielectric (wood, paper, cotton, celotex) and metal (steel wool) materials. Now the cage had become more than an electric and electromagnetic shield - it was on its way to also become a calorimeter, and one that could be made relatively efficient at trapping and concentrating heat. These are the origins of the present cabinet. In essence its construction does not alter the fundamentals once laid out by Reich [5], but makes some improvements that resulted from our experimentation with, and study of, these devices over the years. Thus, for example, the type and thickness of the inner cage, as well as the insulating dielectric, were optimized to produce the desired absorption and attenuation results. The present study permits us, in light of the research that we have previously conducted [6], to present new data on the physiological effect of ORACs, and throw a new light on the operation of these simple converters of ambipolar energy (please see Discussion section).
We have discovered that simple Faraday cages passively operate as energy converters and absorbers of ambipolar (massfree electric) radiation, whose exact spectrum we have identified [7-10]. Whether the ambipolar source is solar, atmospheric or geological ("geothermal"), these cages passively convert incident ambipolar radiation into sensible heat [12-15] (whether kinetic or blackbody) and latent heat [3, 16-17]. This conversion can, in fact, be made very effective, generating enough heat to power a thermal engine with mechanical outputs greater, on a per area basis, than existing photocell technology [17]. Moreover, the devices can be made to output the same amount of mechanical work during nighttime as they do during daytime exposure to solar radiation [17-18].
Years before we understood the functions of the ORAC, we had experimented with cabinets such as those designed by Reich for "medical human application". We observed that, if the weather conditions were anticyclonic, exposure inside the cabinet induced a functional fever in virtually all subjects, as Reich claimed to have observed, and did so as early as after 5 minutes or less of exposure. Over the decades, we accumulated repeated observations, but were never in a position to present them. This is the first report based on such observations, and it focuses on the physiological effect of a particular ORAC that was built for a terminal colon cancer patient and used regularly for nearly 15 years by this patient, as well as tested at various other times with other subjects. The data unequivocally presents evidence for the passive induction of functional hyperthermia in test subjects exposed for 5 to 45 minutes inside ORACs designed for human use, with ΔT values up to 0.9°C, and maximum temperature near the oral ceiling of functional fever (37.5°C). The response plateaus as early as 15 min, but may continue to increase up to 45 min.
The ORAC was built with two traverse full openings on the top and bottom of the front door (respectively 2 and 1 inches) to permit air circulation and eliminate the trapping of exhaled gases. The ORAC cabinet was kept indoors throughout these experiments. Some experiments were conducted with a chair shooter (also 5x double layered) that was placed inside the ORAC, but these are not discussed in the present report. A shooter conduit and funnel built of the same galvanized steel permitted the application of the trapped energy to different segments of the body, as indicated by Reich (results not show in the present report).
Oral temperatures were, in all cases, taken with certified and calibrated medical mercury thermometers, of identical make and provided to the test subjects. Digital thermometers were found to be notoriously unreliable and inconsistent. The test subjects were asked to sit still for 3.5 minutes for each oral temperature reading.
Most human subjects that expose themselves to a properly built ORAC (no copper or aluminum metal parts, no ferromagnetic or other metallic elements on the outer structure, an optimal ferromagnetic and continuous inner cage, an effective thermal insulator with dielectric properties) note a variety of perceptible sensations after some minutes of exposure: tingling on the extremities of limbs or fingers, warmth on the back and palm of hands at 4-8 cm from the walls, heat sensation all over the body and, frequently, sweating after 5 to 15 minutes. Yet, the inside walls of an ORAC cabinet feel cold to the touch (in fact, their temperature is frequently below ambient; see below). Reich also reported skin tanning with prolonged exposure.
2. Systematic induction of functional hyperthermia by exposure of a cancer
patient to a "medical ORAC"
We wanted to investigate whether this 'heat sensation' and sweating reported by most test subjects corresponded to an objective alteration in body temperature and homeothermal control, and what was the magnitude of the elevation, if any, of body temperature above normal with systematic exposure, weather variation and intra-session time-course.
The answer as to whether the phenomenon of 'heat sensation' is merely subjective or also objective, is readily apparent from the results presented in the following three figures.
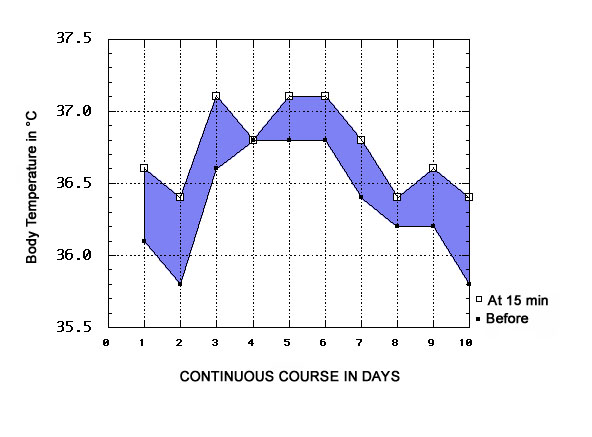
Fig. 1 presents a continuous 10-day course (carried out in Toronto, beginning September 18, 1979) of ORAC exposure during the morning period (09:00 to 10:30). It compares the initial oral temperature taken at t=0 before entering the cabinet, with the oral temperature taken inside the cabinet after 15 minutes of exposure. The physiological ΔT at 15 min is positive in all cases (see also Fig. 2), except for one instance where it is zero (day 4). Careful notes were taken of the weather during this period, which was exceptional in that 8 out of the 10 days were bright, sunny, anti-cyclonic days with either no clouds or only midday cumulus humilis, and the odd cirrus in the morning or at the end of the day. In fact, it was only on the cyclonic day which presented uninterrupted cloud cover that the value of physiological ΔT=0 was registered. On another cyclonic day (day 8), a small positive physiological ΔT=0.2°C was observed.
|
|
| Fig. 1: Body temperatures in the morning, before and after 15 min of ORAC exposure |
The observed positive values of the induced hyperthermia at 15 min exposure ranged from 0.2 to 0.6°C, with a mean ΔT of 0.42°C±SEM 0.05°C (n = 9, see Fig. 2), if we abstract from the null result on day 4. The 15 min ΔT mean for only the anticyclonic days (n=8) is slightly but nonsignificantly higher, at 0.45°C±SEM 0.05°C.
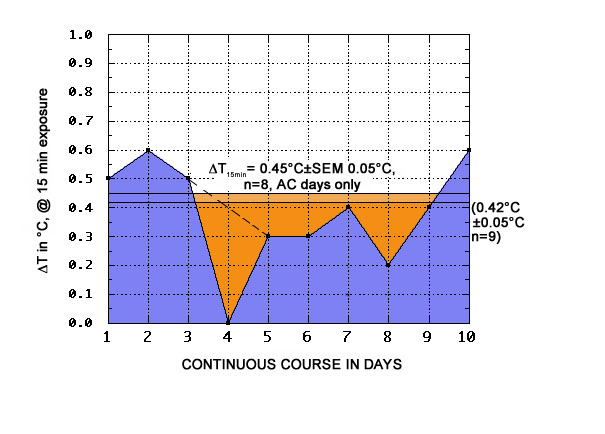
|
|
Fig. 2: ΔT values for body temperatures in the morning, before and after 15 min of ORAC exposure |
The test subject was a 53 year-old male, terminal colon cancer patient diagnosed at the hemorrhagic stage. He had been operated upon 2 weeks before the above 10-day period of ORAC exposure and was, at the time, implanted with a temporary colostomy. He had refused chemotherapy (and had not undergone Cobalt 60 radiotherapy). A month after these tests, he had another surgical intervention to remove the colostomy and religate the colon. The patient underwent no further conventional medical treatment. Throughout the month before the removal of the temporary colostomy, he used the ORAC for irradiation 2 to 3 times daily, on anticyclonic days.
3. ORAC-induced functional hyperthermia is independent of time of day on
anticyclonic days.
Throughout that Winter and Spring, weather and other conditions permitting, the same patient assiduously employed the ORAC cabinet for exposure during periods up to 45 minutes long, 1 to 2 times per day.
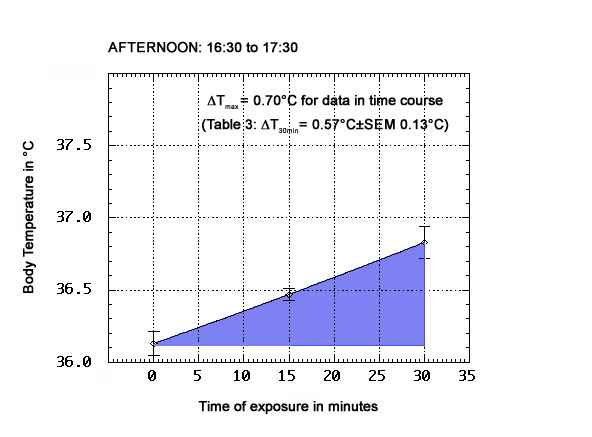
As shown in Tables 1-3 below, the physiological ΔT values obtained for this test subject after 30 min exposure to the ORAC environment on anticyclonic days, and in different periods of the day (morning: 09:00 to 10:00; midday: 12:00 to 13:00; afternoon: 15:00 to 16:00, which is late afternoon in this part of the year, at the latitude of Toronto), are all positive. The top and bottom panels present the means (±SEM) of n=6 groups of observations, with n=8 for the midday panel. ΔT's of up to 0.9°C were observed in every tested period of the day. The morning ΔT mean was 0.58°C±SEM 0.12°C; the midday ΔT mean was 0.59±SEM 0.06°C, and the afternoon ΔT mean was 0.57°C±SEM 0.13°C. A two-tailed Student t test (5df) did not disprove the null hypothesis of no significant difference between these means, or permit its rejection (t=0.056, when comparing morning to afternoon results).
TABLE 1 - Morning period physiological temperatures and ΔT
T @ t=0
(°C)T @ 30 min
(°C)ΔT
(°C)36.8 37.1 0.3 36.4 36.8 0.4 36.2 37.1 0.9 35.9 36.3 0.4 35.6 36.2 0.6 36.2 37.1 0.9
TABLE 2 - Midday period physiological temperatures and ΔT
T @ t=0
(°C)T @ 30 min
(°C)ΔT
(°C)36.5 37.1 0.6 36.7 37.3 0.6 36.3 36.8 0.5 36.0 36.9 0.9 35.8 36.6 0.6 36.2 36.6 0.4 36.9 37.4 0.5 35.8 36.4 0.6
TABLE 3 - Afternoon period physiological temperatures and ΔT
T @ t=0
(°C)T @ 30 min
(°C)ΔT
(°C)36.0 36.6 0.6 36.0 36.6 0.6 36.2 36.9 0.7 36.2 37.1 0.9 36.2 36.8 0.6 37.1 37.2 0.1
Fig. 3 (panels A, B and C) presents results of body temperature readings for
exposure time-courses taken only on anticyclonic days, and at fixed periods
of the day (morning, midday and afternoon). All the results were normalized
to the mean temperature of each daytime period before ORAC exposure, and
their means and standard errors of the mean recomputed. Temperature readings
were taken before the ORAC exposure session, and then at 15 and at 30
minutes of the same session. Collecting data for systematic points was not
easy, since we could not control the weather, nor enforce the times of day
chosen by the patient to employ the ORAC; furthermore, the patient was not
always systematic at recording the temperature before exposure and at the 15
min data point, and had been instructed to end the exposure session whenever
he felt too hot, was sweating too much or simply felt he had had enough. In
sensibly half of the cases, the patient terminated the session at 20 to 25
minutes of exposure. So, the number of instances tested that presented a
full time-course during the same time periods and with the same time-course
points varied, with n=5 for the morning and afternoon periods, and n=3 for
the noon period. All the tested time-courses presented in Fig. 3 took place
on sunny, cloudless days.
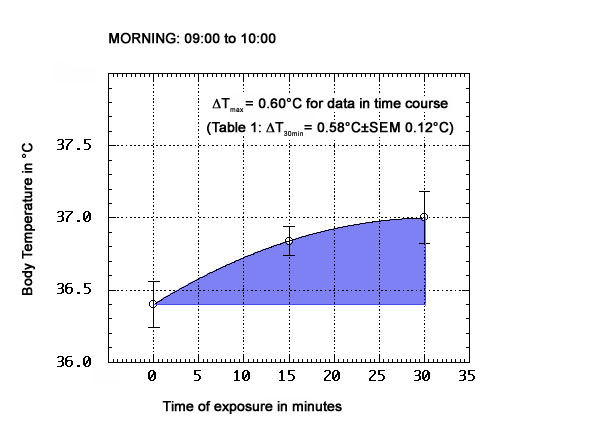
|
| Fig. 3A: Variation of body temperatures with duration of ORAC exposure (morning data) |
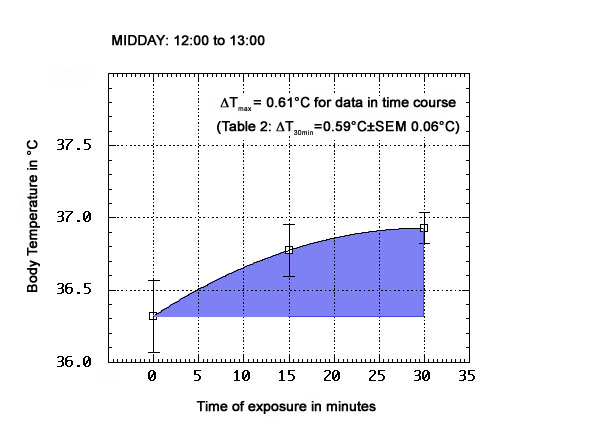
|
| Fig. 3B: Variation of body temperatures with duration of ORAC exposure (midday data) |
|
| Fig. 3C: Variation of body temperatures with duration of ORAC exposure (afternoon data) |
With respect to a comparison of the mean temperature values in each normalized curve of Fig. 3, none of the 30 min means are, statistically speaking, significantly different from the 15 min means; even the afternoon comparison of those means by a two-tailed Student t test fails to reject the null hypothesis (2df, t=1.706, p<0.3). The induction of functional fever by ORAC exposure is evidently a fairly rapid process. In fact, elevation of body temperature can already be detected after 5 minutes of ORAC exposure (see next section). Yet, in all cases, the means clearly rise at 30 min relative to the 15 min means.
4. Intra-session time courses of ORAC exposure for the same test subject
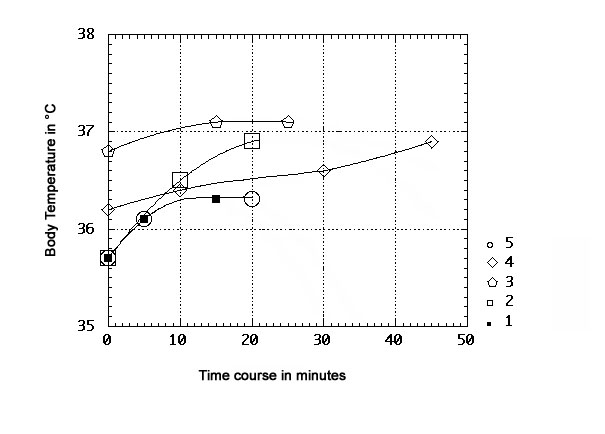
The coefficient of rise of the temperature of the test subject can be seen (Fig. 4) to vary significantly from test to test when comparing selected examples of time-courses from other sessions (irrespective of time of day) containing data points that were not consistently taken at 15 and 30 min, but at other time intervals. This serves as an example of the observed variability in these curves. The temperature during a session may plateau at or near 15 minutes, or instead continue to rise steadily for 45 minutes.
|
|
Fig. 4: Body temperature data from 5 different ORAC sessions, observed at time-points other than the 15- or 30-minute mark. |
The results of Fig. 4 also underline the danger inherent in normalizing the data by the start temperature mean of a given set of time courses, inasmuch as the results suggest that the effect of the ORAC exposure is all the more pronounced when the inactive subject has a temperature closer to the basal temperature (the 'floor') of the normal oral range (note that 3 of the sessions begin at 35.7°C, technically 0.2°C below the conventional 'normal floor'). At starting physiological temperatures closer to the upper level (the 'ceiling') of the normal range, the ORAC effect of elevating body temperature is still detected, but its span - and degree of temperature rise per minute - decrease. Thus, at a 35.7°C starting level, the coefficient of ΔT increase over time is 0.08°C/min for two experiments at 5 min (curves 1 and 5, Fig. 4), and the same for one experiment at 10 min (curve 2, Fig. 4), whereas starting from a 36.2°C level, the coefficient at 10 min is much lower, by 0.06°C/min, at 0.02°C/min (curve 4, Fig. 4). Similarly, at the 35.7°C starting level and at 15 min, the coefficient is 0.04°C/min (curves 1 and 5, Fig. 4), whereas starting 1.1°C higher, at 36.8°C and at 15 min, the coefficient is lower - at 0.02°C/min (curve 3, Fig. 4).
5. Results with other test subjects exposed to the same ORAC
Results from other test subjects exposed to the same ORAC (for control purposes) exhibited, in essence, the same behaviour, as shown in Table 4.
TABLE 4
Subject Activity Status
before exposureWeather Exposure Time
(min)ΔT
(°C)1. TA
35 yr-old maleInactive AC 20 0.65 2. GM
54 yr-old male
w/ permanent
colostomy (*)Inactive
InactiveAC
AC30
250.40
0.503. PN
50 yr-old maleInactive
ActiveAC
AC30
300.55
-0.304. WP
55 yr-old maleInactive C 25 0.20 5. US
58 yr-old maleInactive AC 30 0.50 6. MA
58 yr-old femaleInactive AC 30 0.35 7. AN
54 yr-old femaleInactive
InactiveC
AC30
300.25
0.50
(*) Not due to cancer.
Note that the second result for test subject #3 underlines the importance of
taking the starting temperature when the subject is not in an active state
(see Discussion below). This places a high onus of responsibility on the
researcher who must learn to control the variables properly in order to
consistently observe the physiological thermal effect of ORACs.
6. Operating temperatures of the open ORAC environment
Temperature measurements were carried out in the same ORAC using identical -10 to +50°C calibrated mercury thermometers with a resolution of 0.05°C (for the calibration procedure, see [11]). Measurements were carried out inside the ORAC (in air near the top; on the underside of the top plate and on the walls - by contacting the thermometer bulbs with the metal surfaces of the inner cage), at 3 cm above the top metal plate of the ORAC, and outside (control) the same ORAC, with the door closed and no test subject inside. The results indicated that whereas (1) the air above the top was consistently warmer than the control thermometer by 0.1 to 0.3 °C, and (2) the same could generally be said of the inside air near the top, (3) the underside of the top plate was either at the control air temperature or even cooler, and (4) the walls often cooler than the control.
A typical afternoon time-course of temperatures (corresponding to the 'afternoon' period) taken indoors (with the heating system turned off) - outside (control), inside (near the top of the cage and directly on the underside of the top plate of the cage) and above the same ORAC on a cloudless anticyclonic day - illustrates this behaviour well:
TABLE 5
Control versus Above-Top-of-ORAC temperatures:
Time (min) Control Air Temp Top ORAC Temp ΔT = To-T 16:15 24.6°C 24.7°C +0.10°C 16:45 24.2°C 24.4°C +0.20°C 17:10 23.95°C 24.1°C +0.15°C 17:20 23.9°C 24.05°C +0.15°C 17:30 23.7°C 23.8°C +0.10°C
Control versus Inside (Top) ORAC temperatures:
Time (min) Control Air Temp Inside ORAC Temp ΔT = To-T 16:15 24.6°C 24.8°C +0.20°C 16:45 24.2°C 24.3°C +0.10°C 17:10 23.95°C 23.9°C - 0.05°C 17:20 23.9°C 24.05°C +0.15°C 17:30 23.7°C 23.9°C +0.20°C
Control versus Underside of the ORAC Top Metal Plate temperatures:
Time (min) Control Air Temp Top Underside Temp ΔT = To-T 16:15 24.6°C 24.6°C 0.00°C 16:45 24.2°C 24.2°C 0.00°C 17:10 23.95°C 23.7°C - 0.20°C 17:20 23.9°C 23.9°C 0.00° 17:30 23.7°C 23.8°C +0.10°
The attentive reader of Aetherometry will remark that the positive To-T
values of the first two panels are much lower than those observed in indoors
experiments carried out with 1x (or 2-ply) ORACs or even simple
ferromagnetic Faraday cages [11], and yet the "medical ORAC" has 5 double
layers (10-ply). The reason for this is simple: the "medical ORAC" is an
open cabinet, not a closed box like those experimental devices are; the
"medical ORAC" has two door-wide openings - a 2 inch opening at the top of
the door, and a 1 inch opening at the bottom - which sets up sufficient air
movement inside the box to substantially decrease the temperature difference
(To-T) when compared to closed boxes, even uninsulated ones. This is a very
pertinent fact, in that the physiological temperature elevations observed in
the test subjects cannot be attributed to the modest positive temperature
difference inside the box when compared to the outside (indoors) ambient air
- a temperature difference which is of lower magnitude than the temperature
range of the induced hyperthermia.
The cabinets are not 'distinctly hotter' than the outside, and thus it is not the heat accumulated inside of them which causes the observed functional fever. But even if that were the case, the body does not respond to increases of 0.6°C in ambient temperature with functional fever. A functional fever change of that magnitude would require a substantial elevation of ambient air temperature, one that would have to reach more than 5°C above the physiological mean (desert environment temperatures). Thus, the observed, reproducible functional hyperthermia induced by exposure to an ORAC cabinet lacks any conventionally accepted explanation.
Several conclusions result from our simple investigation of the effect of human irradiation with a "medical ORAC" (Reich's term). These are the facts from the studies above:
2. Fever and functional hyperthermia
Human beings are endotherms, that is, living systems that regulate their internal temperature by endogenous production of heat. Within a temperature range (and qualified by other energy parameters - such as proper nutrition, liquid intake, solar exposure and absence of thermally stressing environments, etc), their internal temperature is fundamentally independent of the temperature of the outside environment, or external sources of heat. Within endotherms, human beings are homeotherms, ie endotherms that maintain a nearly-constant body temperature ('homeothermal control') by employing negative feedback control.
Functional fever is a poorly understood physiological function of homeothermal systems. Body temperature in humans is mostly controlled by the hypothalamus (regulator of autonomic functions), though not exclusively [20]. Thermal regulation of body temperature in homeotherms is achieved by radiative heat loss at the periphery (adrenalin-mediated sympathetic response regulated by the middle hypothalamic nuclei occupying the midpart of the tuber cinereum) and heat production in tissues, particularly liver, brown fat cells (rich in mitochondria), brain and muscles. The heat production is generally a vagotonic response that involves metabolic control and increases the oxygen demand. Once warmed by fever, blood cells, such as monocyte-macrophages (see below), trigger the hypothalamus to dissipate the heat by the sympathetic response: peripheral (cutaneous) dilation, sweating and the shivering mechanism.
Normal body temperature taken orally ranges between 35.9°C and 37.2°C [21] (the rectal and vaginal ranges are 0.3 to 0.5°C higher), varying from individual to individual, with age and time of year, as well as diurnally. Typically, though only as a rule of thumb, body temperature varies from lowest levels in the early morning to highest levels in the late afternoon; this general pattern also applies to pathogenic fever, no matter whether it is sustained, intermittent, remittent or relapsing. The amplitude of the diurnal variation (the circadian range) is 0.6°C [22]. The diurnal range of pathogenic fever is typically much greater than the circadian range, on the order of 1 to 2°C depending on the infectious agent(s), the patient's condition and the type of fever.
Elevations of body temperature can be broadly classified into functional, mechanical and pathogenic. Functional fever, in essence, is any elevation of body temperature which is neither pathogenic nor caused by mechanical injury. Its typical range (magnitude of ΔT: up to 0.5°C) is comparable to that of the circadian range, but may span up to 1.0 °C. In general, this range widens if the functional fever occurs against a lower baseline, say closer to 36°C than to 37°C. Perhaps for this reason, functional fever was considered in the past to define the temperature range of 37.0 to 37.5°C. It is in this sense that an older Merck Manual edition states: "Oral temperatures above 37.0°C (98.6°F) in persons confined to bed [inactive], and above 37.2°C (99.0°F) in persons who are moderately active, usually constitute fever" [23]. Even though in this statement the physiological range of ΔT attributed to activity and detected orally is small (0.2°C), the notion is the same.
Pathogenic fever typically, but not always (for ex. in most cases of Epstein-Barr virus-induced infectious mononucleosis), spans a much greater temperature difference than functional fever, up to 3°C or more in severe diseases. Permanent elevation of body temperature above 41°C may result in brain damage. Above 43°C, heat stroke and death are common.
The basic breakdown of the types of fever, then, is as follows:
1. Functional, if it is caused by
2. Mechanical, if it is caused by:
3. Pathogenic, if it is
Functional fever states are typically unnoticed by the subject. Conversely pathogenic fever states do not normally escape notice, as they are accompanied by other symptoms such as headache, chills, anorexia, nausea, vomiting - and, above 39.5°C, delirium.
It is unclear whether pathogenic fever is or is not advantageous to the patient in infectious and noninfectious conditions [22]. The experimental evidence suggests that in infectious conditions, the febrile state enhances the immune response and thus should not be suppressed (by typical cyclo-oxygenase inhibitors such as acetaminophen [Tylenol] and acetylsalycilic acid [Aspirin]) [22]. In part, the problem harks back to the nature of the circuit inducing the temperature elevation. There are two types of pyrogens that can cause elevation of body temperature: endogenous and exogenous. Endogenous pyrogens (Interleukins 1, 6 and 11; Tumor Necrosis factor, TNF; Interferons; Leukemia Inhibitory factor, LIF; oncostatin M; etc) act by inducing metabolic alterations in the hypothalamic cells in charge of thermal regulation. The main source of endogenous pyrogens are the Il-1 producing macrophages that play a central role in cell-mediated immunity. The principal advantage of the febrile state brought about by an endogenous stimulation is to increase O2 availability by increasing blood supply to affected or infected areas. For every 1°C increase above 37°C, oxygen demand (and consumption, if oxygen is available) increases by 13% [22]. Side effects that may or may not be desirable prominently include increased capillary permeability which, above 38°C, may promote peripheral congestion and edema.
Exogenous pyrogens (or cytokines), on the other hand, also increase body temperature but indirectly, by inducing release of endogenous pyrogens. Here is where, in general, the problem of the usefulness of pathogenic fever for the prognosis of a disease or illness arises, since not all exogenous pyrogens are released by host cells; many are released directly by infectious agents, such as the so-called endotoxin (lipopolysaccharides) of gram-negative bacteria.
Thermal regulation in response to radiant sensible heat appears to involve two different networks of hypothalamic nuclei, one assigned to detecting cold stress (which increases its base frequency of 10 action potential discharges per second with increasing cold) and the other assigned to detect heat stress (which keeps a frequency of 3.7 discharges per second) [24]. However, little is known about how other types of radiation affect the behavior of these two types of thermal detector networks, or other types of networks.
The question of whether febrile states are or not advantageous for the cure of infectious disease is certainly different from the question of the potentially prophylactic employment of ORAC-exposure-induced hyperthermia in normal subjects or in subjects affected with non-infectious illnesses, in particular diseases involving a psychosomatic component (autonomic diseases, likely including non-familial malignancies) or an auto-immune response. We cannot answer this question in the present study; but we remark, in passing, that the colon patient used as the main test subject in this study continued to use this ORAC for 15 years, is still alive, active and in good health at the time of this writing, 27 years after the diagnosis of his illness and the surgical interventions. Proper studies are needed to permit identification of the anticipated beneficial role of ORAC-induced functional hyperthermia in preventing and curing disease.
Accordingly, what is of particular interest to us in this context is the potential medical utilization of induced and controlled functional fever. In this respect, we also note that exposure to solar radiation is thought to cause functional fever (up to 24 hours after exposure) by induction of subcutaneous erythema (followed on subsequent days by tanning), and this is generally attributed to the ultra-violet component of the solar-blackbody spectrum. The fever will be more severe than functional and last significantly longer in cases of severe exposure, such as sunburn and sunstroke. [25] However, there are good reasons to think that solar radiation is not electromagnetic (see below), and that other forms of radiative energy can also induce hyperthermic states. Indeed, exposure to nonlethal doses of ionizing radiation is known to induce febrile states, and their intensity and temperature bear a certain proportionality to the nature and intensity of the radiation. It is also known that medical diathermy can induce functional fever for 6 to 10 hours after application. Of greater interest to us is the possibility that benign ambipolar radiation (massfree electric energy) may also induce hyperthermic states, because of its likely direct biological involvement in supplementing thermal control and regulation of body temperature.
3. A biophysical process involved in the thermal control and regulation of
endotherms?
Presently, the discussion will focus on three facts borne out by prior studies that we conducted:
- That the nature of solar radiation is not electromagnetic but ambipolar [8-9].
- That the interaction of ambipolar radiation with metallic and dielectric materials gives rise to both in situ production of blackbody photons and the capture of latent heat, by molecules that absorb or attenuate ambipolar radiation [8, 11, 19].
- Following our identification of the ambipolar spectrum [8], we distinguished between two subspectra - the OR and DOR ranges of ambipolar energy that have opposing biological effects, respectively, beneficial and detrimental - and identified their electric energy (79.4keV) and frequency (2.18*1018 sec-1) boundaries [8].
These facts indicate that exposure of the human body to solar radiation leads to the in situ cutaneous and subcutaneous conversion of the incident radiation into photons and latent heat. However, this is not the only physical process for the absorption of solar ambipolar radiation. For ambipolar radiation may be partially absorbed and still pass through the tissues in attenuated form. Moreover, ambipolar energy (in the OR subspectrum, see [8]) may well play a normal role as a biological form of energy [20]. Lastly, solar radiation is not the only source of ambipolar energy, since atmospheric gases and petrogeological sources can also emit ambipolar radiation. In fact, the discovery of the roles played by latent heat and ambipolar energy in diverse physical and biological systems forced us to provide a new foundation for a theory of energy systems [20] that also corrects some fundamental mistakes in various approaches to thermodynamics.
We have previously reported on the results of our formal study which suggested that other biophysical mechanisms were involved in the regulation of the temperature control of endotherms. In particular, several lines of evidence suggest that thermal production and regulation is not simply metabolic, but may involve a biological ambipolar field capable of affecting the ordinary electric fields of the body and their surface potentials, as well as indirectly generating blackbody heat (sensible heat in photon form). In this context,
1) we have proposed a new (aetherometric), linear temperature scale and a consistent aetherometric value for the Boltzmann constant [11, 13];
2) we demonstrated that if a blackbody heat bath with a temperature of 37°C were to be indirectly produced by an ambipolar field - or directly released by an electron plasma - it would require a field voltage of 513V, or be capable of imparting to electrons a kinetic energy of 513 eV [8-9, 26];
3) the observation that even in the most efficient metabolism (characteristic of beta-state aerobes such as human beings), chemical energy only accounts for 1/4th of the energy required to reduce oxygen to water and generate ATP [27] led us to suggest that it is ambipolar energy (in the OR subspectrum) which contributes the missing energy [27];
4) we formally demonstrated that pigments such as protoporphyrin IX (required by hemoglobin) function as ambipolar OR energy absorbers (28.28 keV for hemoglobin) that generate blackbody heat as their output [28-29];
5) we identified in the chromatin-DNA structural complex, a self-inductive step-down autotransformer for ambipolar radiation, capable of absorbing ambipolar OR energy of 34 keV and down-converting its voltage to 499 volts, with a temperature of nearly 30°C [26].
Finally, these lines of evidence raised the question of whether biological systems, in particular endotherms, deploy an ambipolar field (passively, actively, or in both manners) that may indirectly supplement internal heat production (such a field could also be present in ectotherms [30]).
The only recognized source of biological heat is metabolic, and it invokes only biochemical processes, especially those that employ the reserves of brown fat cells. But our present inquiry leads us to ask whether there is also a biophysical process which is directly involved in thermal regulation, even if mediated at various points by specific chemical substrates. The preceding leads us to propose that:
1) endotherms tend to regulate their internal temperature near the temperature of the ambipolar field released by the cellular nucleus (ie the DNA complex in the cellular nucleus);
2) the DNA complex is tuned to receive solar ambipolar radiation of 34 keV and emit an an ambipolar field near 500V;
3) the universality of the protoporphyrin ring IX as a biological ambipolar antenna and an electromagnetic emitter suggests that aside from oxygen transport and delivery, blood - or more specifically, red blood cells - also act as absorbers and accumulators of ambipolar energy (tuned to receive ambipolar radiation of 28.2 keV) [29] that release blackbody photons (at the red to infra-red boundary).
Thus the erythemic response to solar exposure is more than a mechanical flooding of the peripheral tissues and blood vessels caused by thermal dilation of the capillaries; because of the blood hemoglobin content, the body 'knows' that the red blood cells are elements tuned to receive precisely the solar-sourced ambipolar energy required to form oxygen from ozone and release infrared heat, so it mobilizes them to absorb that ambipolar radiation and generate heat. In the process, transient skin erythema and induction of functional fever take place, as long as the exposure to the solar radiation is moderate. Thus - we suggest - absorption of ambipolar radiation, attenuated or not, and above all from the solar source, defines a precise biophysical process in endotherms that can directly contribute and affect heat production and regulation.
Further, we suggest that melanocytes constitute further proof of this "unconscious knowledge" on the part of the body. Though not part of the present study, the role of melanin in absorbing both blackbody photons and ambipolar radiation, and transducing their energy into heat (sensible or latent) is parallel to the biophysical role of the globins in the blood, and the functional reason why melanocytes are developed from basal stem cells of the epidermis, and remain in locations near the basement membrane of the epidermis. There is a whole plant-becoming of animals, going on just underneath the skin, and the erythemic response associated with functional fever is a dynamic part of it, perhaps the main process for the dissipation of energy acquired radiatively, whether electromagnetic, ambipolar or latent heat.
4. ORAC-exposure-induced transient hyperthermia
Faraday invented the Faraday cage and observed its electric insulating properties, and Reich invented the ORAC and observed its thermal (anomalous heat) and electroscopic properties (anomalously slow rate of leakage) [3, 11]. Reich was also the first to report on the biological and biophysical effects of human exposure to an ORAC environment. Unfortunately his methodology was not always the best or most systematic, and the induced hyperthermia was one of those poorly demonstrated results of 'orgone irradiation' that received recognition in medical Reichian circles without a real scientific investigation. On this topic Reich made one remark:
"This fact [ie of "orgonotic excitation" by the superimposition of two energy systems, the ORAC and the human being inside of it] becomes perceptible to the user of the orgone energy accumulator through feelings of stronger prickling, warmth, relaxation, reddening of the face, and, objectively, through increased body temperature (0.5 to 1.5 °F). The body temperature should be measured before and during the irradiation (not after the sitting)". [31].
That is exactly what we did in all the observations reported above, and in other instances, and what Reich (or his group of young medical doctors) perhaps should have done, if for no other reason than to counteract the FDA assault that destroyed his research process.
Prior to the present report, there is only one other report on the subject of the ORAC-induced hyperthermia that these authors are aware of. This was part of a joint (?) dissertation in the Department of Psychology at the University of Marburg, Germany [32]. There, however, the authors only present histograms (equivalent to our Table 4, but unfortunately with the data normalized, so that the real temperatures were not reported), with no statistics (either multi-session for each test subject, or for the subject population) and without a single intra-session time-course, or a day sequence. The same study also failed to explain the negative result it obtained with one subject (seemingly, an instance of induced hypothermia). Despite these severe limitation, it had the virtue of showing that the hyperthermia was not observed upon exposure to a control box devoid of metal, when an ORAC and the control box were compared with a double-blind protocol.
In our present study, we found that the range of the observed induced functional hyperthermia was slightly higher than claimed by Reich (0.2 to 0.9°C, or up to nearly 2°F), and was independent of the chosen subject and thus an objective fact, but that, in general, it could only be observed on anticyclonic days. It also increases with the exposure time-course, typically plateauing at 30 minutes of exposure.
How do aetherometric lines of inquiry and evidence discussed above relate to the results of the present study? To us, they suggest the following explanation to the observed transient hyperthermia induced by exposure to an ORAC cabinet:
1) That what causes it is neither blackbody heat nor a hot environment (sensible heat), since the cabinet is dark, placed indoors, constitutes an electromagnetic shield, and is not distinctly warm, let alone hot, compared to the ambient air outside of it;
2) That its cause can be: either absorption of attenuated ambipolar energy; or absorption of the latent heat generated by the conversion of that ambipolar radiation at the metallic walls of the inner enclosure; or both.
In ORAC enclosures optimized for maximal direct solar exposure and the generation of round-the-clock maximal temperature differences (To-T of up to 35°C or more), the amount of latent heat generated from the conversion of incident solar ambipolar radiation can be as high as 230 times, or more, the amount of sensible heat (including blackbody heat) generated inside the same enclosure [33]. We have also shown that if the cabinet is not earth-grounded, ambipolar radiation can still traverse it, though in attenuated form [8]. With ambipolar field voltages of 50 kV, near the solar mode, we were able to charge circuits placed inside ORAC cages of 1/4th the thickness of the "medical ORAC" employed in this study, with potentials of at least 90 to 120V, with a vacuum tube receiver [8]. Much greater "filtered" voltages were detected with simple metallic pick-ups. It is also worth mentioning that the antenna sensor of a biofield meter of our design and capable of detecting and measuring biological ambipolar radiation, induces skin sensations of heat and tingling on the palm and the back of the hand when the test subject's hand is within the distance where the meter registers the ambipolar field of the subject. Effectively, the test subject feels the blackbody heat resulting from the dissipation of their ambipolar field on the electrons trapped on the surface of the antenna sensor. This is analogous to what is experienced by ORAC-exposed test subjects, and to us it suggests two conclusions:
1) The sensation of physiological heat, the sweating and the hyperthermia induced by ORAC exposure must be attributed, in pars or in toto, to the absorption of attenuated ambipolar energy by the human body. The main source of the attenuated ambipolar energy is the Sun.
2) The sensation of tingling or warmth near the walls of the ORAC must be differently attributed to the interaction of the ambipolar energy field of the human body (the ambipolar "aura") with the metallic walls of the cabinet.
5. A regulatory role for the pineal gland in the biological response to
ambipolar radiation?
The pineal gland is a neuroendocrine organ that, in mammals, develops from both mesoderm and neural crest cells. The pineal "complex" is found in all vertebrates. In "evolutionarily lower" vertebrates, where the complex can be found subepidermally on the median dorsal aspect of the head, the main activity of the pineal gland (epiphysis) has been suggested to be photoreception (a "third eye"), but the fact remains that even in these species the eye-like structure remains diminutive and buried under the skin. In "higher" vertebrates the gland "evolutionarily" migrated inward to the center of the brain and is thought to have a "vestigial" photoreceptor "function".
In mammals, the pineal gland is one of the key components of the neuroendocrine system. Within it, neural (norepinephrine synthesized by the sympathetic fibers innervating the pineal body) and hormonal (indoleamines such as melatonin or its precursor, serotonin; and peptides or proteins) components interact to regulate the operation of the gland. The two major functions of the gland are the control and modulation of the circadian and circannual biological rhythms, and thermoregulation of the internal environment.
The great interest and mystery of the functional role of the pineal body is that, while it has been shown to control the seasonal regression of reproductive organs in photoperiodic species (eg the golden hamster [34]), the circadian function of the gland is in fact independent of light and photo-control. The circadian rhythmicity is currently considered to be endogenous or innate, which is just a way of saying that its functioning is not known. Rat pups raised in continuous darkness or continuous light retained their circadian rhythms, even if these were observed to drift or to be susceptible to entrainment (as by the rhythm of a nursing foster mother) [35-36]. Similar findings have been experimentally documented in other species, avian in particular [37].
The conclusion was inevitable: the biological clock of serotonin is "generated independent of environmental light-dark schedules" [38]. This has led to the notion that the pineal was once a photoreceptor but has now ceased to respond to direct illumination, even if (chick) pinealocytes grown in tissue culture respond to constant light by suppressing the nighttime increase in N-acetyl serotonin synthesis [39]. However, precisely retina cells do not exhibit that behavior, thus differing from pinealocytes in the light-response [40]. Chronobiology is often claimed to be a new science, but it is hard to envision how it can move forward when it is unable to understand how the pineal gland controls circadian rhythms, other than by invoking "endogenous" factors that are suggested to be genetic, or take recourse to entrainments caused by a variety of factors (nursing mother rhythm, displacement of photic stimuli, "nonphotic clues", temperature entrainment, etc). Studies that support an autoregulatory clockwork model - where the protein products of the suprachiasmatic nuclei of the hypothalamus clock genes (Period and Cryptochrome) regulate the expression of other genes - have failed to identify the precise trigger and input signals that feed the clockwork.
Our explicit suggestion, in this context, and one that could be easily proven with an experimental arrangement, is that the pineal body is not really a photoreceptor organ, but an ambipolar sensor organ. In line with the aetherometric theory of light as being produced indirectly and locally in the form of photons by the propagation of ambipolar radiant energy, cellular photoreceptors may have specialized in two distinct directions -
1) those that receive both photic stimuli in the outer environment (optical vision, or ordinary vision) and ambipolar radiation from without - radiation capable of triggering on the surface of special cells a local production of photons; this would be the role of retinal cells and largely why they differentiated into two different types. And
2) those which only receive ambipolar radiation, the pinealocytes, and specialized therefore in forming "dark receptors"; these could recede to the innermost cavity of the body to receive attenuated ambipolar radiation in the upper frequency or energy range of the ambipolar spectrum (high voltage OR and DOR). Electronic coupling of pinealocytes is in fact suspected to control the flow of the synthetic output of pinealocytes [41]. Moreover, pineal modulation of the thermal control exerted by the hypothalamus may further suggest that the same pinealocytes may also receive or sense the ambipolar radiation coming from the internal environment (via the blood system), and this may well be the evolutionary sense of their close coupling to blood capillaries in the perivascular spaces.
This proposed model admits entirely to the possibility of photon production inside the two subcellular processes of pinealocytes: in both cases the receptors would absorb ambipolar radiation and release photons in situ, but there is no need to invoke any reception of external light (photic) signals. Moreover, our model also suggests that the intraparenchymal and perivascular processes of pinealocytes may well have differentiated to detect, respectively, the attenuated external ambipolar radiation, and the internal ambipolar radiation. Or, it may be that the perivascular process is capable of detecting directly the ambipolar-radiation-induced photon production from, for example, hemoglobin (the lizard Sphenodon has an intracranial pineal gland equipped with a lens and pigment-containing pinealocytes). Either way, this would explain why the function of the pineal body is effectively independent of external light cycles, and why its cells are capable of being "entrained" by temperature manipulation of an in vitro simulated internal environment [42].
In light of this aetherometric model, any increase of the internal temperature detected by pinealocytes (either as internally produced ambipolar radiation or as photon production, as from hemoglobin), would be compared with the reception of attenuated ambipolar radiation from the external environment, and if the latter matched or more than matched the former, it would trigger a functional hyperthermic entrainment of the body "thermostat" setting that fit the amount of irradiation. Such an upward thermostat drift would be exemplified by the ORAC-induced hyperthermia. Its thermostat drift is generally greater than that observed in exercise-induced functional hyperthermia, in which internal ambipolar radiation would increase but no absorption of outside ambipolar radiation would correlate with it. Similarly, direct skin exposure to solar radiation can easily trigger greater thermostat drifts than those induced by exercise. The "vagotonic balance" to the sympathetic innervation of the pineal would, in essence, be provided by the ambipolar stimuli. Finally, the pineal independence from light variation and the light/dark cycle, and its capacity to retain a circadian clock and resynchronize it, would be simply pineal responses to the diurnal variation of the intensity of solar ambipolar radiation reaching the Earth's surface. The endogenous clock would thus turn out to be but a comparative ambipolar clock.
1. Williams, P (1965) "Michael Faraday", Da Capo Press, NY. NY, p. 291.
2. Correa, P & Correa, A (1997) "Non-equivalence between work performed by charge against gravity and the electric energy of the same 'charge gas'", Akronos Publishing, Concord, Canada, ABRI monograph AS2-01.
3. Correa, P & Correa, A (1998, 2001) "Comparative study of the variation in the spontaneous discharge rate of atmospheric electroscopes and electroscopes placed within 'orgone accumulators'", Akronos Publishing, Concord, Canada, ABRI monograph AS2-06.
4. Correa, P, Correa, A & Askanas, M (2002) "Atmospheric electricity, latent heat, and ambipolar radiation: a new view of geophysics and meteorology, challenging the primacy of ionization theory" Akronos Publishing, Concord, Canada, ABRI monograph AS2-27.
5. Reich, W (1951) "The Orgone energy accumulator - its scientific and medical use", Orgone Institute, Orgonon, ME.
6. Correa, P & Correa, A (2003) "Experimental Aetherometry", Vol.s I, IIA & IIB, Akronos Publishing, University of Toronto Press, Concord, Canada.
7. Correa, P & Correa, A (1999) "Aetherometric treatment of the energy radiation output by Tesla coils (3): 1° massfree electric-and-magnetic waves, 2° massbound capacito-inductive waves and 3° electromagnetic waves", Akronos Publishing, Concord, Canada, ABRI monograph AS2-16.
8. Correa, P & Correa, A (1999) "The indirect 'ORgone effect' of Tesla radiation: solar spectra for ambipolar Aether and Blackbody radiation", Akronos Publishing, Concord, Canada, ABRI monograph AS2-17A.
9. Correa, P & Correa, A (2000) "Determination of the OR and DOR energies, frequencies and wavelengths driving the atmospheric allotropic cycle of oxygen, ozone and water", Akronos Publishing, Concord, Canada, ABRI monograph AS2-17B.
10. Correa, P & Correa, A (2000) "The cosmic background microwave radiation as evidence for cosmological creation of electrons with minimum kinetic energy and for a minimum of cosmic ambipolar massfree energy", Akronos Publishing, Concord, Canada, ABRI monograph AS2-17C.
11. Correa, P & Correa, A (1998, 2001) "The thermal anomaly in ORACs and 'the Reich-Einstein Experiment': implications for blackbody theory", Akronos Publishing, Concord, Canada, ABRI monograph AS2-05.
12. Correa PN & Correa AN (2001) "The reproducible thermal anomaly of 'the Reich-Einstein Experiment' under limit conditions", Infinite Energy, 37:12.
13. Correa, P & Correa, A (1998, 2001) "Decoding the thermal and nonthermal equivalents of the org as a unit of aether energy", Akronos Publishing, Concord, Canada, ABRI monograph AS2-07.
14. Correa PN & Correa AN (2002) "A modified orgone accumulator (HYBORAC) as a drive for a low Delta-T Stirling engine - Part I", Akronos Publishing, Concord, Canada, monograph AS2-25.
15. Correa PN & Correa AN (2002) "A modified orgone accumulator (Complete HYBORAC) as a nighttime drive for a low Delta-T Stirling engine - Part II", Akronos Publishing, Concord, Canada, ABRI monograph AS2-26.
16. Correa PN & Correa AN (2002) "A modified orgone accumulator (Complete HYBORAC) as a nighttime drive for a low Delta-T Stirling engine - Part II", Infinite Energy, 42:41.
17. Correa, P & Correa, A (2003) "Around the clock free power from improved HYBORACs driving low delta T gamma Stirling engines - Part IV", Akronos Publishing, Concord, Canada, ABRI monograph AS2-32.
18. Correa PN & Correa AN (2004) "Free Power around-the-clock", an ABRI/Akronos Production DVD, Akronos Publishing, Concord, Canada.
19. Correa, P & Correa, A (1999) "(Re-)Examination of the energy radiation output by Tesla coils: (1) Experimental determination of its dual nature", Akronos Publishing, ABRI monograph AS2-13.
20. Correa, P & Correa, A (2004) "Nanometric functions of bioenergy", Akronos Publishing, University of Toronto Press, Concord, Canada.
21. Chatton, MJ (1980) "General Symptoms", in "Current Medical Diagnosis & Treatment", ed. M. Krupp & M. Chatton, Lange medical Publications, Los Altos, CA, p. 1.
22. The Merck Manual, 17th edition, 1999, p. 1093.
23. The Merck Manual, 13th edition, 1997, p. 4.
24. Tromp, S (1980) "Biometeorology", Heyden, London, England, p. 65.
25. Induction of functional fever in UV tanning beds, though rather poorly studied, may constitute evidence that blackbody photons alone can elicit the effect, even if the present report suggests that such induction does occur with other forms of energy and in the absence of significant blackbody photons.
26. Correa, P & Correa, A (2004) "Nanometric functions of bioenergy", op. cit., p. 301.
27. Ibidem, pp. 259 and following.
28. Ibidem, pp. 266-269.
29. Correa, P & Correa, A (2002) "Fundamental measurement of biological energies 1: overview of bioenergetic investigations", Akronos Publishing, Concord, Canada, ABRI monograph AS2-28.
30. Without us claiming by the present study that such a biophysical process as we suggest also applies to ectotherms, it is apparent that the main source of energy tapped by ectotherms (which depend on the environment to maintain their body temperature) is solar radiation (ambipolar) or its byproducts (sensible and latent heats).
31. Reich, W (1951) "The Orgone energy accumulator", op. cit., p. 27.
32. Muschenig, S & Gebauer, R (1986) "The psychophysiological effects of the Reich orgone energy accumulator", University of Marburg, Germany.
33. Correa, P & Correa, A (2003) ABRI monograph AS2-32, op. cit., p. 37.
34. Ablation of the pineal gland leads to suppression of the seasonal regression, see Weiss, L (1983) "Histology - cell and tissue biology", Elsevier Science Publishing Co., NY, NY, p. 1080.
35. Deguchi, T (1975) "Ontogenesis of a biological clock for serotonin: acetyl coenzyme A N-acetyltransferase in pineal gland of rat", PNAS (USA), 72(7):2814.
36. Takahashi, K & Deguchi, T (1983) "Entrainment of the circadian rhythms of blinded infant rats by nursing mothers", Physiol Behav, 31:373.
37. Zawilska, J et al (2006) "Daily oscillation in melatonin synthesis in the Turkey pineal gland and retina:diurnal and circadian rhythms", Chronobiol, 23:341.
38. Deguchi, T (1978) "Ontogenesis of circadian rhythm of melatonin synthesis in pineal gland of rat", J Neural TransmSuppl, 13:115.
39. Deguchi, T (1981) "Rhodopsin-like photosensitivity of isolated chicken pineal gland", Nature, 290:706.
40. Zawilska, J & Wawrocka, M (1993) "Chick retina and pineal gland differentially respond to constant light and darkness: in vivo studies on serotonin N-acetyltransferase (NAT) activity and melatonin content", Neurosci Lett, 153:21.
41. Weiss, L (1983) op. cit., p. 1085.
42. Barrett, R & Takahashi, J (1995) "Temperature compensation and temperature entrainment of the chick pineal cell circadian clock", J Neurosci, 15:5681.